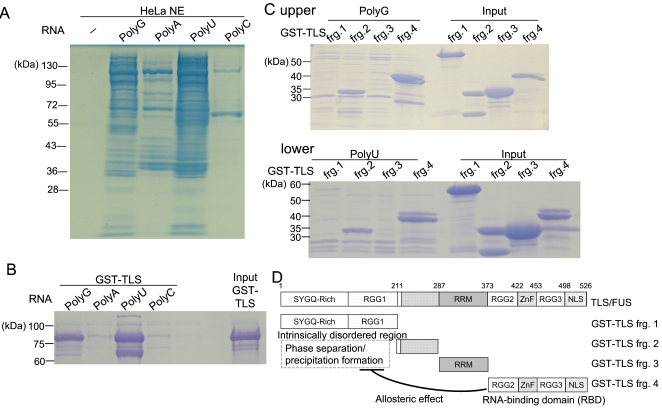
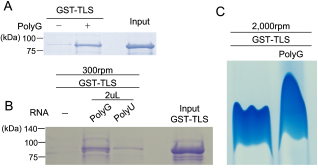
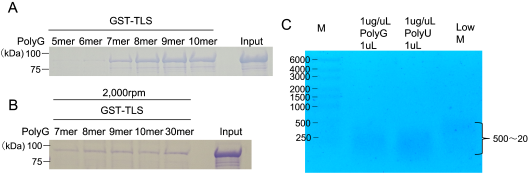
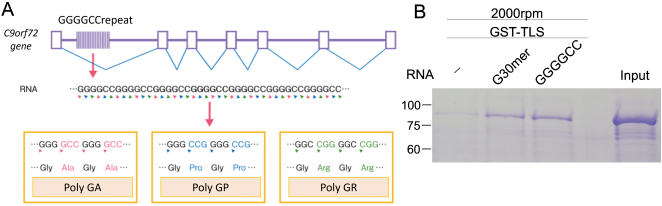
Translocated in Liposarcoma (TLS), also known as Fused in Sarcoma (FUS), is a multifunctional RNA-binding protein implicated in neurodegenerative diseases due to its tendency to aggregate. While mutations in TLS are linked to familial amyotrophic lateral sclerosis (ALS), approximately 90% of ALS cases are sporadic with no known genetic mutations. In these instances, pathological aggregation of wild-type TLS is believed to play a critical role, although the molecular triggers remain elusive. RNA is known to modulate TLS phase separation, but the features that drive RNA-induced precipitation are poorly understood. Here, we report that synthetic PolyG RNA robustly induces both phase separation and irreversible precipitation of recombinant TLS in vitro. This effect is concentration-dependent and strongly influenced by RNA sequence composition. Specifically, guanine-rich RNAs such as PolyG promote aggregation, whereas uridine-rich RNAs fail to induce precipitation and may even inhibit it. These findings suggest a selective interaction between TLS and G-rich RNA sequences. Notably, the resulting TLS-RNA complexes undergo precipitation in a manner distinct from classical liquid-liquid phase separation, highlighting a unique mechanism of RNA-induced protein misfolding. Through detailed molecular biological and biochemical analyses, we further demonstrate that PolyG-induced condensates transition into solid-like aggregates over time. Our results uncover a previously uncharacterized pathway of RNA-mediated TLS aggregation and suggest that guanine-rich RNAs may contribute to pathological protein misfolding in neurodegenerative disease contexts.
| Published in | Biomedical Sciences (Volume 11, Issue 4) |
| DOI | 10.11648/j.bs.20251104.11 |
| Page(s) | 70-77 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
TLS/FUS, ALS, Phase Separation, Poly G, RNA, IDR
ALS | Amyotrophic Lateral Sclerosis |
BISOX | Biotinylated Isoxazole |
CamKIIa | Ca/calmodulin-dependent Protein Kinase Type II Subunit Alpha |
CBB | Coomassie Brilliant Blue |
CBP | CREB-binding Protein |
CNS | Central Nervous System |
DBPs | DNA-Binding Proteins |
FTD | Frontotemporal Dementia |
FTLD | Frontotemporal Lobar Degeneration |
G4 | G-Quadruplex |
GR | Glucocorticoid Receptor |
HAT | Histone Acetyltransferase |
IDRs | Intrinsically Disordered Regions |
iPS cells | Induced Pluripotent Stem Cells |
lncRNA | Long Noncoding RNA |
NLS | Nuclear Localization Signal |
pncRNA-D | Promoter-Associated ncRNA-D |
Poly G | Polyguanylic Acid |
RBPs | RNA-Binding Proteins |
RRM | RNA Recognition Motif |
SDS-PAGE | SDS-Polyacrylamide Gel Electrophoresis |
| [1] | Mao YS, Zhang B, Spector DL. RNA: a bridge between nuclear structure and function. Trends Genet. 2011; 27(8): 295–306. |
| [2] | Fox AH, Lamond AI. Paraspeckles. Cold Spring Harb Perspect Biol. 2010; 2(7): a000687. |
| [3] | Sawyer IA, Dundr M. Nuclear bodies: built to boost. Chromosoma. 2017; 126(1): 1–14. |
| [4] | Milicevic K, Rankovic B, Andjus PR, Bataveljic D, Milovanovic D. Phase separation in neurodegeneration. Front Cell Dev Biol. 2022; 10: 840256. |
| [5] | Patel A, Malinovska L, Saha S, Wang J, Alberti S, Krishnan Y, Hyman AA. ATP as a biological hydrotrope. Science. 2017; 356(6339): 753–756. |
| [6] | Posey AE, Ruff KM, Harmon TS, Crick SL, Li A, Diamond MI, Pappu RV. Profilin reduces aggregation and phase separation of huntingtin N-terminal fragments. eLife. 2018; 7: e36330. |
| [7] | Wright PE, Dyson HJ. Intrinsically disordered proteins in cellular signalling and regulation. Nat Rev Mol Cell Biol. 2015; 16(1): 18–29. |
| [8] | van der Lee R, Buljan M, Lang B, et al. Classification of intrinsically disordered regions and proteins. Chem Rev. 2014; 114(13): 6589–6631. |
| [9] | Uversky VN. Intrinsically disordered proteins in overcrowded milieu: membrane-less organelles, phase separation, and intrinsic disorder. Front Phys. 2019; 7: 10. |
| [10] | Sun Z, Diaz Z, Fang X, et al. Molecular determinants and genetic modifiers of aggregation and toxicity for the ALS disease protein FUS/TLS. PLoS Biol. 2011; 9(4): e1000614. |
| [11] | Neumann M, et al. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and ALS. Science. 2006; 314(5796): 130–133. |
| [12] | Mackenzie IR, et al. Pathological TDP-43 distinguishes sporadic ALS from ALS with SOD1 mutations. Lancet Neurol. 2010; 9(10): 995–1007. |
| [13] | Mariani D, et al. Transcriptomic landscape of G-rich RNAs in human neurons. Nucleic Acids Res. 2024; 52(21): 13269–13289. |
| [14] | Ueda N, Yoneda R, Kurokawa R. TLS/FUS domain analysis in RNA binding and aggregation. Biomed Sci. 2024; 10(2): 45–52. |
| [15] | Cui W, Yoneda R, Ueda N, Kurokawa R. Arginine methylation of TLS inhibits its binding to long noncoding RNA, abrogating TLS-mediated repression of CBP/p300 activity. J Biol Chem. 2018; 293(28): 10937–10948. |
| [16] | Hamad N, Yoneda R, So M, Kurokawa R, Nagata T, Katahira M. Non-coding RNA suppresses FUS aggregation caused by mechanistic shear stress on pipetting in a sequence-dependent manner. Sci Rep. 2021; 11(1): 9523. |
| [17] | Simone R, et al. G-quadruplexes: emerging roles in neurodegenerative diseases. Trends Mol Med. 2015; 21(8): 513–525. |
| [18] | Fay MM, et al. RNA G-quadruplexes are globally unfolded in eukaryotic cells and depleted in bacteria. J Mol Biol. 2017; 429(14): 2127–2147. |
| [19] | Kharel P, et al. RNA G-quadruplexes: emerging insights into regulatory roles and therapeutic potential. Nucleic Acids Res. 2020; 48(22): 12534–12555. |
| [20] | Yamazaki T, et al. Functional domains of TLS/FUS in phase separation and RNA binding. Mol Cell. 2018; 70(6): 1038–1053.e7. |
| [21] | Statello L, Guo CJ, Chen LL, Huarte M. Gene regulation by long non-coding RNAs and its implications for disease. Nat Rev Mol Cell Biol. 2021; 22(2): 96–118. |
| [22] | Lin Y, Protter DSW, Rosen MK, Parker R. Formation and maturation of phase-separated liquid droplets by RNA-binding proteins. eLife. 2015; 4: e06807. |
| [23] | Aoki M. Clinical features of ALS in Japan. Rinsho Shinkeigaku. 2013; 53(11): 1080–1083. |
| [24] | Vance C, Rogelj B, Hortobagyi T, et al. Mutations in FUS, an RNA processing protein, cause familial ALS. Nat Genet. 2009; 41(4): 409–414. |
| [25] | Kwiatkowski TJ Jr, Bosco DA, Leclerc AL, et al. Mutations in the FUS/TLS gene on chromosome 16 cause familial ALS. Nat Genet. 2009; 41(4): 439–443. |
| [26] | Bosco DA, Lemay N, Ko HK, et al. Mutant FUS proteins that cause ALS incorporate into stress granules. Nat Neurosci. 2010; 13(5): 545–552. |
| [27] | Renton AE, Majounie E, Waite A, et al. A hexanucleotide repeat expansion in C9ORF72 is the cause of chromosome 9p21-linked ALS-FTD. Neuron. 2011; 72(2): 257–268. |
| [28] | Mori K, Weng SM, Arzberger T, et al. The C9ORF72 GGGGCC repeat is translated into aggregating dipeptide-repeat proteins in FTLD/ALS. Science. 2013; 339(6125): 1335–1338. |
| [29] | Fujino Y, Ueyama M, Ishiguro T, et al. GGGGCC repeat RNA promotes TLS aggregation in ALS models. eLife. 2023; 12: e84338. |
| [30] | Ash PE, Bieniek KF, Gendron TF, et al. Unconventional translation of C9ORF72 GGGGCC expansion generates neurotoxic DPRs. Neuron. 2013; 77(4): 639–646. |
| [31] | Smith RA, et al. Methods for analyzing RNA–protein interactions in phase separation. Methods Mol Biol. 2019; 2040: 115–126. |
| [32] | Kato M, et al. Cell-free formation of RNA granules: low complexity sequence domains form dynamic fibers within hydrogels. Cell. 2012; 149(4): 753–767. |
| [33] | Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970; 227(5259): 680–685. |
| [34] | Sugimoto N, et al. Thermodynamic characterization of G-quadruplex formation by guanine-rich RNA. Molecules. 2020; 25(21): 4936. |
| [35] | Zhong S, et al. G-rich RNA transcripts promote phase separation of FUS through G-quadruplex formation. Acta Neuropathol. 2024; 148: 21. |
| [36] | Yamashita R, et al. G-intron database: analysis of G-rich intronic sequences. Genome Res. 2005; 15(5): 589–601. |
| [37] | Oyoshi T, et al. C-rich complementary sequences suppress G-quadruplex-mediated aggregation of RNA-binding proteins. Chem Pharm Bull. 2025; 73(10): 938–943. |
| [38] | Sato K, et al. Duplex formation between G-rich RNA and C-rich DNA prevents phase separation of FUS. Brief Bioinform. 2021; 22(6): bbab321. |
| [39] | Liu X, et al. Molecular chaperones modulate RNA-induced phase separation of FUS. Molecules. 2023; 28(15): 5801. |
| [40] | Teng Y, et al. Chemical modulation of RNA–protein condensates: implications for neurodegenerative disease. Bull Chem Soc Jpn. 2025; 98(10): 1123–1134. |
| [41] | Haberman N, et al. Insights into RNA–protein interactions in neurodegeneration. BMC Biol. 2024; 22: 254. |
| [42] | Wu Y, et al. RNA-mediated phase separation of FUS in C9orf72 ALS. Neuron. 2024; 112(3): 456–472. |
| [43] | The Lancet Neurol. Functional aggregation in neurobiology: a double-edged sword. Lancet Neurol. 2025; 24(10): 1002–1015. |
| [44] | Hervás R, et al. Molecular basis of memory-related amyloid-like aggregates in Drosophila. Science. 2020; 367(6483): 1230–1234. |
APA Style
Ueda, N., Yoneda, R., Kurokawa, R. (2025). PolyG RNA Induces Phase Separation and Precipitation of TLS/FUS. Biomedical Sciences, 11(4), 70-77. https://doi.org/10.11648/j.bs.20251104.11
ACS Style
Ueda, N.; Yoneda, R.; Kurokawa, R. PolyG RNA Induces Phase Separation and Precipitation of TLS/FUS. Biomed. Sci. 2025, 11(4), 70-77. doi: 10.11648/j.bs.20251104.11
AMA Style
Ueda N, Yoneda R, Kurokawa R. PolyG RNA Induces Phase Separation and Precipitation of TLS/FUS. Biomed Sci. 2025;11(4):70-77. doi: 10.11648/j.bs.20251104.11
@article{10.11648/j.bs.20251104.11,
author = {Naomi Ueda and Ryoma Yoneda and Riki Kurokawa},
title = {PolyG RNA Induces Phase Separation and Precipitation of TLS/FUS},
journal = {Biomedical Sciences},
volume = {11},
number = {4},
pages = {70-77},
doi = {10.11648/j.bs.20251104.11},
url = {https://doi.org/10.11648/j.bs.20251104.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.bs.20251104.11},
abstract = {Translocated in Liposarcoma (TLS), also known as Fused in Sarcoma (FUS), is a multifunctional RNA-binding protein implicated in neurodegenerative diseases due to its tendency to aggregate. While mutations in TLS are linked to familial amyotrophic lateral sclerosis (ALS), approximately 90% of ALS cases are sporadic with no known genetic mutations. In these instances, pathological aggregation of wild-type TLS is believed to play a critical role, although the molecular triggers remain elusive. RNA is known to modulate TLS phase separation, but the features that drive RNA-induced precipitation are poorly understood. Here, we report that synthetic PolyG RNA robustly induces both phase separation and irreversible precipitation of recombinant TLS in vitro. This effect is concentration-dependent and strongly influenced by RNA sequence composition. Specifically, guanine-rich RNAs such as PolyG promote aggregation, whereas uridine-rich RNAs fail to induce precipitation and may even inhibit it. These findings suggest a selective interaction between TLS and G-rich RNA sequences. Notably, the resulting TLS-RNA complexes undergo precipitation in a manner distinct from classical liquid-liquid phase separation, highlighting a unique mechanism of RNA-induced protein misfolding. Through detailed molecular biological and biochemical analyses, we further demonstrate that PolyG-induced condensates transition into solid-like aggregates over time. Our results uncover a previously uncharacterized pathway of RNA-mediated TLS aggregation and suggest that guanine-rich RNAs may contribute to pathological protein misfolding in neurodegenerative disease contexts.},
year = {2025}
}
TY - JOUR T1 - PolyG RNA Induces Phase Separation and Precipitation of TLS/FUS AU - Naomi Ueda AU - Ryoma Yoneda AU - Riki Kurokawa Y1 - 2025/12/17 PY - 2025 N1 - https://doi.org/10.11648/j.bs.20251104.11 DO - 10.11648/j.bs.20251104.11 T2 - Biomedical Sciences JF - Biomedical Sciences JO - Biomedical Sciences SP - 70 EP - 77 PB - Science Publishing Group SN - 2575-3932 UR - https://doi.org/10.11648/j.bs.20251104.11 AB - Translocated in Liposarcoma (TLS), also known as Fused in Sarcoma (FUS), is a multifunctional RNA-binding protein implicated in neurodegenerative diseases due to its tendency to aggregate. While mutations in TLS are linked to familial amyotrophic lateral sclerosis (ALS), approximately 90% of ALS cases are sporadic with no known genetic mutations. In these instances, pathological aggregation of wild-type TLS is believed to play a critical role, although the molecular triggers remain elusive. RNA is known to modulate TLS phase separation, but the features that drive RNA-induced precipitation are poorly understood. Here, we report that synthetic PolyG RNA robustly induces both phase separation and irreversible precipitation of recombinant TLS in vitro. This effect is concentration-dependent and strongly influenced by RNA sequence composition. Specifically, guanine-rich RNAs such as PolyG promote aggregation, whereas uridine-rich RNAs fail to induce precipitation and may even inhibit it. These findings suggest a selective interaction between TLS and G-rich RNA sequences. Notably, the resulting TLS-RNA complexes undergo precipitation in a manner distinct from classical liquid-liquid phase separation, highlighting a unique mechanism of RNA-induced protein misfolding. Through detailed molecular biological and biochemical analyses, we further demonstrate that PolyG-induced condensates transition into solid-like aggregates over time. Our results uncover a previously uncharacterized pathway of RNA-mediated TLS aggregation and suggest that guanine-rich RNAs may contribute to pathological protein misfolding in neurodegenerative disease contexts. VL - 11 IS - 4 ER -
Division of Biomedical Sciences, Saitama Medical University, Saitama, Japan
Division of Biomedical Sciences, Saitama Medical University, Saitama, Japan
Division of Biomedical Sciences, Saitama Medical University, Saitama, Japan